
RICARDO MOTTA PINTO COELHO
RICARDO MOTTA PINTO COELHO
PROJETO 7
Introdução
Em Ecologia, existem basicamente três tipos de modelos matemáticos: a) os modelos estatísticos também chamados de modelos empíricos; b) os modelos determinísticos, também chamados de modelos dinâmicos e os modelos mistos que englobam características dos dois primeiros modelos (Hakanson & Peters, 1995). Os modelos empíricos baseiam-se em regressões simples, múltiplas, lineares ou não. A principal característica desse tipo de modelo é a busca de relações entre as variáveis de estudo que a princípio NÃO são conhecidas. Por isso mesmo, o modelo é exploratório ou empírico. A segunda categoria envolve o uso de equações diferenciais onde os processos a serem modelos são conhecidos, a priori. O modelo determinístico requer o uso de parâmetros pré-determinados em equações que definem processos precisos. É, sem dúvida uma categoria mais elegante e precisa de modelos ecológicos. Eles têm uma definição teórica que, no entanto, nem sempre reflete a realidade. Por outro lado, os modelos empíricos são pura expressão da realidade, mas qual realidade é essa? A frase de extraída de Hankanson & Peters (1995) reflete - de modo emblemático - a dualidade entre esses dois tipos de modelos: "in dynamic models, one may say that the scientific knowledge is used to set the process, but guesses are used to set the rates. In empirical models, science is used to get the constants (rates) but guesses are made about the process".
O objetivo de todo modelo é explicar o funcionamento de um determinado sistema. Esse sistema pode ser tão grande quanto a biosfera ou uma simples população de ciliados em um frasco de culturas. A premissa básica de todo modelo, por mais complexo que possa ser, é que trata-se de um método limitado de representar a natureza. Assim, todo modelo precisa de ser testado, de ser validado ou, mais importante ainda, de ser falseado! Em essência, a principal característica de um ecólogo modelista consiste em aprimorar o seu modelo, num contínuo processo de busca de erros, para a sua depuração, através da proposição de um novo modelo. Pode-se mesmo dizer o objetivo mais importante é verificar sob quais condições o modelo NÃO funciona, ou seja, verificar e comprovar quais a limitações dos modelos. No entanto, esse é um processo extremamente salutar pois vai permitindo ao ecólogo uma visão muito aprimorada dos processos e estruturas que governam o funcionamento do ecossistema.
Os ecossistemas são extremamente complexos de serem estudados considerando todos os seus
componentes. Uma das formas de se entender essa complexidade, consiste em modelilzar o
comportamento de certas variáveis complexas, que são tomadas como variáveis de estado nos
modelos. Em limnologia, essas variáveis são basicamente
aquelas influenciadas por
um elenco de diversos fatores bióticos e abitóticos. Exemplos dessas
variáveis seriam o pH, o fósforo (dissolvido ou total) ou a profundidade
do desaparecimento do
disco Secchi (transparência). O esquema abaixo nos dá uma idéia da multiplicididade de fatores
relacionados com a disponibilidade de fósforo na água de um lago ou reservatório.
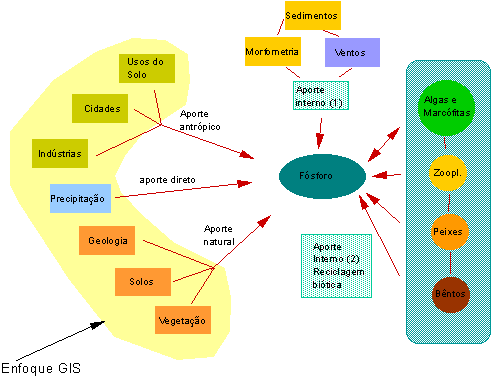
Muitos poderão perguntar nesse ponto: porque estudar algo tão complicado como o fósforo em um rio, lago, laguna ou reservatório? A Ecologia é uma ciência que ainda carece ainda de princípio fundamentais (ver livro Fundamento de Ecologia, Introdução, pg. 12). No entanto, um dos princípios mais universais dessa ciência é a relação encontrada pelo pequisador suíço- canadense Vollenweinder (1976) entre o fósforo total e a clorofila-a. Apesar das inúmeras variações existentes, essa relação empírica é consistente sendo válida em todos os ecossistemas aquáticos, sem restrições. Trata-se de uma simples regressão linear que ilustra a dependência da biomassa algal em relação ao fósforo, um nutriente essencial frequentemente limitante para a produção primária. Para ter maiores informações sobre os fatores limitantes à produção primária nos ecossistemas recomendamos uma leitura do capítulo 18, pg. 175, do livro Fundamentos em Ecologia.
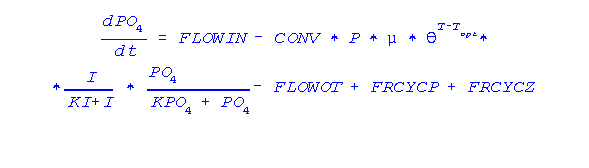
O modelo que usaremos, a seguir, relaciona o fósforo dissolvido na água do lago ao processo da produção primária, ao metabolismo do zooplâncton e de seus predadores. O modelo pode ser definido pelo conjunto de equações diferenciais abaixo. A primeira variável de estado é a concentração de fósforo dissolvido na água. Ela depende da taxa de aporte (FLOWIN) que pode ser tanto interno (via sedimentos) e externo (tributários), da assimilação do fitoplâncton (que depende de vários fatores, tais como radiação e temperatura), da excreção das algas (FRCYCP), do zooplâncton (FRCYCZ) bem como da taxa de saída do sistema (FLOWOT).
A segunda variável é a biomassa de algas fitoplanctônicas (P). Essa variável depende da taxa de
crescimento do fitoplâncton que por sua vez, depende da própria biomassa (P) no instante t0, da
taxa de crescimento (µ), da taxa de radiação luminosa (I), da concentração de fósforo (PO4), da
temperatura da água (T), da taxa de sedimentação (SED), da taxa de herbivoria (GRAZ) bem
como da taxa de diluição do meio (FOUTP).
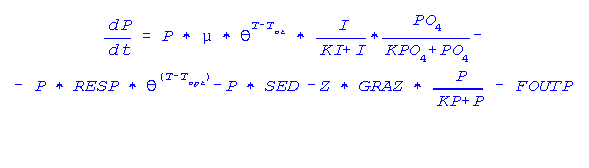
A terceira variável de estado é a biomassa de organismos zooplanctônicos (Z). Ela depende da biomassa do zooplâncton (Z) em t0, da taxa de herbivoria (GRAZ ), da concentração do fitoplâncton (P), da eficiência de conversão energética (ASS). Como principais fatores de perda temos o gasto energético via respiração, que depende fortemente da temperatura (T), a predação que depende da taxa de predação (CARN) e a saída do sistema por diluição, wash out (FOUTZ).

A quarta variável de estado é a biomassa de carnívoros, ou consumidores de zooplâncton (C) no sistema. Ela depende da concentração da peixes zooplanctívoros (CARN), da biomassa do próprio zooplâncton (Z), da eficiência de assimilação (ASSCARN). Como fatores de perda temos as perdas metabólicas (respiração) que depende da temperatura (T) e a remoção via diluição ( wash out ).

Para uma visão mais detalhada dessas equações acima
recomendamos uma leitura do capítulo
25, pg. 233, do livro Fundamentos em Ecologia.
Simulações Realizadas
Em nossos estudos iniciais sobre modelização da comunidade planctônica, estivemos preocupados em adaptar o modelo a condições "tropicais". Após revisão de literatura, consulta ao nosso banco de dados sobre a represa da Pampulha onde pudemos mensurar várias taxas tais como a respiração do zooplâncton e a excreção de nutrientes do zooplâncton e exaustivas simulações propomos, na tabela abaixo, parâmetros para simular condições tropicais e condições temperadas. Os parâmetros em azul são os mesmos tanto para ambas as situações. Os parâmetros em vermelho são mais adequados aos organismos das zonas tropicais.

Para uma discussão mais aprofundada do significado desses parâmetros, consultar o capítulo 25, pgs. 236-238, do livro Fundamentos de Ecologia. Consultar também o capítulo 21, pgs. 199-204, onde publicamos algumas das taxas relacionadas ao metabolismo do zooplâncton em ambientes temperados (L. Constança, Alemanha) e tropicais (represa da Pampulha).
O objetivo das simulações a seguir foi o de estudar as respostas de uma comunidade planctônica de um reservatório tropical ao fenômeno da eutrofização. O modelo abaixo está baseado numa nova versão do mesmo programa apresentado no capítulo 25, do Livro Fundamentos em Ecologia. Nessa nova versão, incluímos uma nova variável de estado, ou seja, os predadores de zooplâncton (C). Portanto, o modelo agora ficou mais completo, suportando uma nova gama de diferentes simulações.
Apresentaremos, a seguir, três simulações realizadas. Nós iremos alterar três diferentes
parâmetros. A taxa de entrada, a de fósforo (FLOWIN), o tempo de retenção (TIMERET) e o
nível de predadores de zooplâncton (CARN). A tabela abaixo
nos dá os valores iniciais e finais de cada
um desses parâmetros nas três simualções realizadas:

As simulações levaram em consideração um período equivalente a 5 anos (1825 dias). Em todas as simulações, em cada ano, usamos valores crescentes de aporte de fósforo para simular um aumento na eutrofização. Na simulação A, temos apenas os resultados do avanço da eutrofização. Nas duas outras simulações (B e C), iremos estudar os efeitos de dois tipos de manejos clássicos usados na recuperação da qualidade de ambientes (reservatórios) eutrofizados: variação do tempo de retenção (simulação B) e biomanipulação (simulação C):
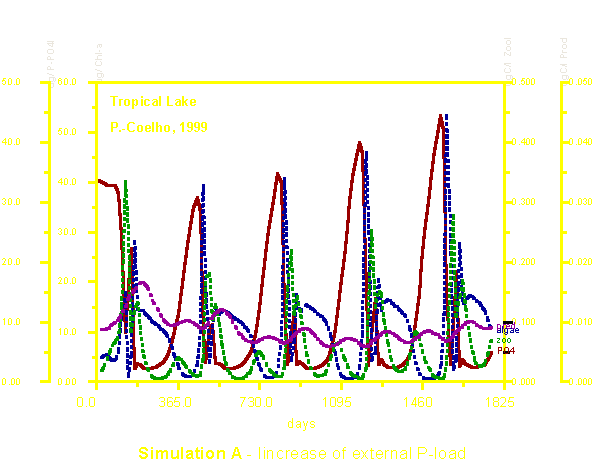
Na simulação A, temos os resultados do modelo para um caso simples de eutrofização causada pelo aumento das médias anuais do aporte de fósforo. Observar que a biomassa algal respondeu de modo claro ao aumento na oferta de nutrientes. O zooplâncton herbívoro também aumentou em densidade mas não pudemos notar qualquer aumento claro no quarto nível trófico, ou seja, predadores de zooplâncton.
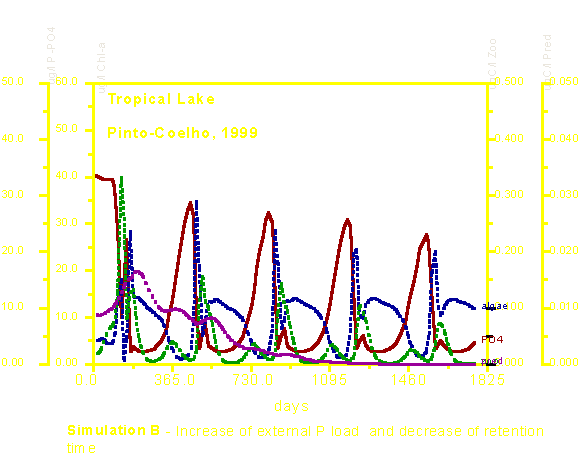
Na simulação B, a situação fica mais complexa, pois o aumento do aporte externo de fósforo é contrabalançado pela diminuição do tempo de retenção da água no sistema (reservatório). Dessa maneira, para uma mesma entrada de nutrientes a resposta em termos de aumento de biomassa algal foi mais discreta. Essa simulação comprova que a gestão de um reservatório (através do manejo das comportas, por exemplo) pode auxiliar para uma melhoria da qualidade da água. Observar que este tipo de estratégia causa ainda grandes impactos na comunidade do zooplâncton (linhas verdes) que fica praticamente impedido de se reproduzir no sistema e portanto decresce de modo agudo em termos de densidade e biomassa. Obviamente o mesmo efeito é visto no quarto nível trófico.
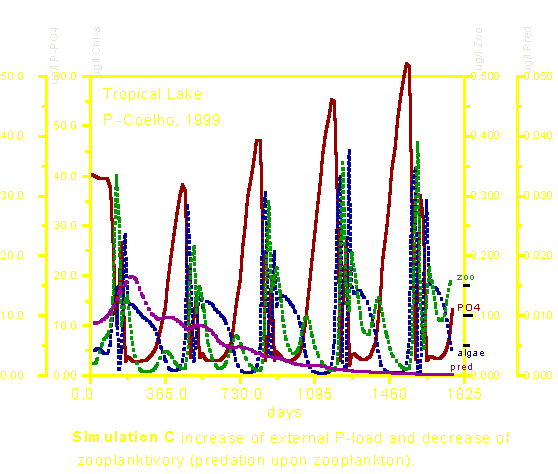
A simulação C é, sem dúvida alguma, a mais interessante delas. Ela demonstra que a diminuição da predação no zooplâncton causa profundos impactos em todos os níveis tróficos ( efeito top down ). Apesar do aporte de fósforo ter sido exatamente o mesmo nas três simulações, vemos aqui que o acréscimo de biomassa de algas é muito discreto. Como diminuir a predação sobre o zooplâncton? Isso pode ser feito basicamente diminuindo-se a população de peixes zooplanctívoros através da adição de peixes piscívoros. Um fato interessante nessa simulação, refere-se aos valores de fósforo solúvel encontrados na água que ficaram mais elevados do que na simulação A mas dada a elevada eficiência da cadeia trófica., os nutrientes não ficam "intrabiotizados" e não geram "capitalização" da biomassa de algas. A qualidade de água permanece adequada apesar do elevado aporte de um elemento limitante. Essa simulação ilustra a potencialidade do "efeito cascata trófica" , ou seja, os efeitos de "topo de cadeia" (top down effects). Os efeitos da predação na estrutura das comunidades são discutidos em detalhes no capítulo 11, pgs. 122-124, do livro Fundamentos em Ecologia.
A tabela abaixo nos dá os valores médios e a amplitude observada para cada uma das quatro variáveis de estado usadas no modelo (fósforo dissolvido, clorofila-a, zooplâncton e predadores do zooplâncton). Os valores médios de clorofila-a foram ligeiramente menores nas simulações B e C. Observar, porém, que a amplitude foi reduzida nessas simulações. Isso pode ser entendido que eventos tais como waterblooms (florescimentos massais de algas) que causam uma grande degradação na qualidade de água ficariam mais limitados, por exemplo. No nível do zooplâncton, a simulação B causou sensível redução nos valores médios enquanto que a simulação C, ao contrário, causou aumento na densidade e biomassa desses organismos. A simulação C, evidententemente causou uma diminuição dos predadores do zooplâncton graças ao aumento de peixes piscívoros.

Programa completo
Aqui você pode ter acesso (download) ao programa completo (em linguagem Turbo Pascal,
DOS) que gerou as simulações acima. Grave esse arquivo em um diretório
C:\TP de seu disco rígido.
Programa DYNAMICS.EXE
Para poder usar a rotina gráfica você vai ter ainda que instalar o seguinte aplicativo no diretório C:\TP de seu disco rígido:
Usando o Windows Explorer, por exemplo, grave os arquivos acima, no diretório C:\TP (tem que ser exatamente com esse nome). Depois, na barra de tarefas do (Win95 em diante), clicar em iniciar, executar, procurar (localizar o arquivo dynamics.exe no diretório C:\TP) e clicar OK. Se tudo der certo, a tela do programa DYNAMICS.PAS irá aparecer na tela de seu computador.
O programa deve ser rodado através do acionamento sequencial das seguintes rotinas (passos):
1) teclar "j" para iniciar o programa para condições tropicais;
2) teclar "a" para visualizar a lista dos parâmetros selecionados;
3) teclar "c" para realizar a simulação A;
4) teclar "g" para visualilzar o gráfico na tela (essa rotina exige drive gráfico);
5) teclar "h" para visualizar gráfico das variáveis de força (essa rotina exige drive gráfico);
6) teclar "s" para selecionar a forma de saída dos resultados;
7) teclar de "1" a "7" para imprimir os resultados;
8) teclar "m" para mudar o status (TRUE ou FALSE) das variáveis
booleanas que controlam as simulações B e C.
Para a simulação B,
a variável TimeRet_Simula_Bol deve ter o valor alterado TRUE e para a simulação
C, a variável Carn_Simula_Bol deve ter o valor alterado para TRUE. Para voltar aos
valores originais basta teclar "j", novamente. Em caso de dúvidas entre em
contato conosco.
Literatura:
Hakanson, L. & R.H. Peters. 1995. Predictive Limnology. SPB Academic Publishing, Amsterdam, The Netherlands, ISBN 90-5103-104-1, 464 p.
Vollenweider, R. 1976. Advances in defining critical loading levels for phosphorus in lake eutrophication. Mem. Ist. Ital. Idrobioll. 33:53-83.

Página criada em 27 de junho de 2001 e atualizada em 31/10/2016
Prof. Ricardo Motta Pinto Coelho
Dépt. Sciences Bbiologiques
Université de Montréal, UdM, Montréal, Québec.
E-mail: rmpc@icb.ufmg.br